domingo, 14 de mayo de 2017
viernes, 21 de abril de 2017
domingo, 12 de marzo de 2017
REFLEXIÓN
LO QUE
APRENDIMOS.
Como ya lo mencionamos antes
en la célula se pueden distinguir dos etapas la interfáse y
la división.
En este tema pudimos aprender que
el núcleo interfásico es el centro de control pues este contiene la mayor parte
de ADN; es el órgano más voluminoso de la célula
eucariota podemos decir que su constitución varía a lo largo de la
vida celular
En la interfáse la célula crece y
se puede observar la forma y la estructura del núcleo y la división, durante la
cual la membrana que la rodea el núcleo desaparece con los componentes del
núcleo se mezcla con el citoplasma.
Su forma puede ser elíptica,
esférica y lobulada.
Su tamaño va relacionada con el
tamaño del citoplasma ya que si su volumen crece el del núcleo también lo hace,
su posición puede ser el el centro de la célula pero también puede desplazarse
lentamente contra la pared celular.
También pudimos
observas partes de la nucleo interfacico
Como la
cromatina es el estado morfológico en el que se encuentra e ADN y la membrana
nuclear se trata de una de las estructura celulares con mayor organización y
complejidad. El cromosoma es la estructura altamente organizada y formada por
el ADN que asu vez por las proteínas ya que tienen la mayor parte genética de
un individuo.
EL NÚCLEO INTERFÁSICO.
En eucariontes el ADN, que
contiene los genes, está separado del citoplasma por la envoltura nuclear. Esto
tiene importantes consecuencias, porque el proceso de transcripción de ARN, a
partir del ADN, está físicamente separado del sitio de la síntesis proteica,
que tiene lugar en el citoplasma.
Los precursores de los ARN
mensajeros, transcritos a partir del ADN en el núcleo, con frecuencia
experimentan un amplio procesamiento, con escisión de segmentos y en empalme de
otros, antes de producirse una molécula de ARN mensajero (ARNm) citoplasmático
maduro. Sólo los ARNm completamente maduros salen a través de la envoltura
nuclear.
En procariontes la maquinaria
de la síntesis proteica no está separada del ADN por una membrana nuclear, y el
ARN copiado a partir de los genes está disponible de inmediato para su
traducción en proteína, Esto produce un tipo muy diferente de regulación
genética.

El microscopio óptico reveló
que el núcleo interfásico está rodeado por una envoltura definida, aun cuando
no fue posible determinar con certeza sus componentes. Mediante el uso de
diversos colorantes básicos se halló en los núcleos interfásicos, que la
cromatina puede encontrarse en estado condensado (heterocromatina) o en forma
dispersa (eucromatina). Además se pueden observar uno o varios cuerpos,
llamados nucléolos. El nucléolo tiene una alta concentración de ARN y
proteínas, no está rodeado por membranas. También hay grandes conglomerados de
cromatina unidos a la parte externa del nucléolo. El resto del núcleo se halla
ocupado por el nucleoplasma en apariencia no estructurado, que presenta
numerosos gránulos densos, algunos de los cuales contienen ARN.
El nucleoplasma es amorfo
cuando se lo observa con el microscopio electrónico, pero probablemente tiene
una estructura subyacente. Los núcleos aislados extraídos por métodos
bioquímicos muy drásticos (como solución salina 1 molar, diversos detergentes y
extensa digestión del ADNasa y ARNasa) retienen todavía algunas de las
estructuras observadas morfológicamente en los cortes para microscopio
electrónico.
ENVOLTURA NUCLEAR
La
envoltura nuclear consta de dos membranas que separan el contenido nuclear del
citoplasma circundante. Las dos membranas de la envoltura se interrumpen en
algunos puntos formando poros nucleares, de tal forma que el interior del
núcleo se comunica con el citoplasma celular. Los poros nucleares presentan
canales acuosos que permiten el paso de sustancias del interior de núcleo hacia
el citoplasma y viceversa, pero el proceso es altamente selectivo, permitiendo
solamente el paso a moléculas específicas.

FIGURA 2- ESTRUCTURA DEL NÚCLEO INTERFÁSICO (MEMBRANAS).
El poro nuclear en realidad es una estructura altamente elaborada, denominada complejo del poro nuclear, compuesta de más de 100 proteínas diferentes, ordenadas con una simetría octagonal. Las moléculas pequeñas (5 kDa o menos) difunden en forma prácticamente libre, pero las proteínas de gran tamaño necesitan contar con una señal de localización nuclear, que generalmente consiste en una corta secuencia de aminoácidos (de 4 a 8).
El
proceso de entrada de una proteína destinada al núcleo necesita que otra
proteína citosólica ("receptor nuclear de importación") llamada
nucleoporina se una a la señal de localización nuclear y requiere además de la
energía que proporciona la hidrólisis de una molécula de trifosfato de
guanidina (GTP). Esto provoca la dilatación del poro y permite el pasaje de la
proteína. La salida de las subunidades ribosómicas fabricadas en el nucléolo y
el ARN también dependerá de un sistema de transporte activo mediado por señales
de exportación nuclear.

FIGURA 3- CAPA DEL NÚCLEO INTERFÁSICO.
Unida
al interior de la membrana nuclear se encuentra una capa de proteínas
específicas que al parecer funcionan como esqueleto del núcleo (lámina nuclear)
y que tiene un papel importante en la desorganización y reorganización de las
membranas nucleares al comienzo y al fin de la división celular,
respectivamente. La lámina nuclear es un enrejado de subunidades proteicas, del
tipo de los filamentos intermedios que han sido vistos al tratar el cito
esqueleto (proteínas fibrilares) y que, como todas las proteínas nucleares
lleva una “señal de transporte nuclear” que los dirige desde el citosol, donde
son sintetizados, hacia el núcleo.

FIGURA 4- PROCESO DE LA DIVISIÓN NUCLEAR
Cuando
el núcleo se desensambla durante la mitosis, la lámina nuclear se depolimeriza
por fosforilación. Al mismo tiempo la membrana nuclear se desarma en vesículas
membranosas, que van adheridas a las proteínas fosforiladas (fracciones de la
lámina nuclear). En la telofase temprana (una de las fases finales de la
división celular) se produce la defosforilación de las proteínas y las
vesículas se repolarizan alrededor de cada cromosoma. En la telofase tardía las
vesículas se reúnen y reconstituyen la envoltura nuclear de cada célula hija,
que activamente reimporta a las proteínas que llevan la señal de transporte
nuclear y recompone los poros nucleares.
LA CROMATINA
El ADN es el principal
componente genético de la célula y el que lleva la información codificada de
una célula a otra y de un organismo a otro. El ADN no se halla libre sino un
complejo, la llamada ¨cromatina¨
La
paradoja del valor c

Todos los núcleos de un individuo tienen una cantidad constante de ADN. Esto surgió que el ADN contenía la información genética se demostró que durante la diferencia de los distintos tejidos somáticos no hay perdida de información y esto se logró coloreando el ADN por la reacción de Feulgen y midiendo la cantidad de colorante en núcleos aislados. Todas las células de un organismo contienen la misma cantidad de ADN (2C), siempre que sean diploides. Los gametos son haploides y tienen la mitad de ADN (1C), algunos tejidos como el hígado, contienen algunas células que son poliploides con núcleos que contienen más ADN.
Cada
especie tiene un contenido de ADN característico, que contienen todos los individuos de esa
misma especie y, por eso se le denomina valor C. En los eucariontes hay grandes
variaciones del contenido de ADN, aunque este siempre es mayor que en los
procariontes. Los eucariontes interiores contienen generalmente menos ADN. Los
vertebrados poseen mayor contenido de ADN (cerca de 3pg) por lo general unas
700 veces más que la E.coli.
Considerando
que un picogramo de ADN equivale a 31 cm de éste y que el contenido total por
célula está en una sola molécula, puede calcularse, sobre base teóricas,
longitudes de 174 cm de ADN en los 46 cromosomas humanos ha sido determinado
citofotométricamente, de estas mediciones surgió la posible proporcionalidad
entre la longitud del cromosoma y su contenido en ADN.
La cromatina es un complejo de ADN e
Histonas
La
cromatina puede aisiarse separando los núcleos y tratándolos y tratándolos con
soluciones hipotónicas. En estas condiciones aparecen como suspensión
gelatinosas que contienen ADN, ARN, proteínas básicas – las histonas – y
proteínas no histónicas, mas acídicas. Desde comienzos de la década de 1960
sabíamos que el contenido de ARN y de proteínas no histónicas varía en
diferentes células pero que las histonas se encuentran siempre en una
relación de peso con el AND.
Las
proteínas no histónicas son muy heterogéneas y varían en distintas células.
Entre ella se incluyen: las ARN polimerasas, las ADN polimerasas y diversas
reguladoras. Son relativamente pequeñas y básicas que contienen entre 10 y 10 %
de los aminoácidos básicos: arginina o lisina. Siendo básicas las histónicas,
se unen fuertemente al ADN que es ácido. Las histonas H2A, H2B,
H3 y H4 son semejantes en diversas especies y se
encuentran entre las proteínas más diversas más conservadas que se conocen.
Estas cuatro histonas se hallan en cantidades equimoleculares (dos de cada una
por 200 pares de ADN).
La histona H1, no se conserva entre
especies y hasta tiene formas específicas para diversos tejidos. Hay solo una
molécula de H1, por cada 200 pares de bases, además, esta histona se
une en forma más laxa, es dable extraerla con soluciones salinas débiles, y
pueden perderse con facilidad durante la preparación bioquímica de la
cromatina. La H1 está relacionada con el plegamiento de cromatina.
Los extendidos de cromatina permiten revelas una estructura
en forma de cuentas de collar
Al
observar extendidos de cromatina se encontró una estructura repetitiva en forma
de cuentas de collar de 10 mm de diámetro, los nucleosomas, conectados entre sí
por un filamento de ADN.
Esta
disposición se comprueba en casi todas las cromatinas observadas, sin embargo
no representa la verdadera estructura, sino un artificio como resultado de la
pérdida de la histona H1.
El Nucleosoma es un actámero de histonas con 200 pares de
bases de ADN
Se
digiere la cromatina como una enzima llamada nucleasas microcócica, se puede
cortar el ADN en segmentos de diferente tamaño, los que son todos múltiplos de
200 pares de bases (pb). Así hay segmentos de 200, 400, 600, 800, etc. En otros
estudios se observó que las histonas H3 y H4 tienden a
asociarse formando tetrámeros de dos moléculas cada una.
Como
las cuatro histonas H2 A, H2 B, H3, H4, se hallan cantidades
equimoleculares, Roger Kornberg, en 1974, propuso un modelo de nucleosoma, en
el cual las cuatro histonas se disponen formando un octámero con 200 pares de
bases de AN. Los octámeros están en contacto íntimo (como sucede en los
neuclosomas de la fibra 10nm) y el ADN se enrola en la periferia de nucleosoma.
Los
neuclosomas adyacentes están unidos por el ADN de conexión, o puente que está
más expuesto a la enzima. Una breve digestión da origen a los nucleosomas, que
están formados por un octámero de histona H, 200 pares de base de ADN y una
molécula de histona H1. El nucleosoma es una partícula aplanada en
forma de disco de 11 nm de diámetro y 5,7 nm de altura. El ADN describe dos
giros completos alrededor de los octámeros de histona y estas dos vueltas se
estabilizan con una molécula de H1. La digestión muy fuerte genera
una partícula o núcleo del nucleosoma (core)
de 146 pares de bases que ha perdido el ADN de conexión.
El ADN entra y sale del nucleosoma en sitios
próximos entre su y las dos vueltas de ADN son estabilizadas o podríamos decir
“selladas”, por la misma histona H1. La cromatina que no tiene H1origina
el aspecto en cuentas de un collar en el cual el ADN entra y sale de los
neuclosomas al azar. La histona H1 también puede interactuar con la
H1 de otros nucleosomas adyacentes y esto lleva a un mayor
plegamiento de la fibra.
El
largo de ADN por nucleosoma puede variar en distintos tejidos. Sin embargo, la
variación solo se produce en el puente de ADN y no en la parte central del
neuclosoma, que siempre contiene una cantidad constante de ADN.
Cuando
los nucleosomas forman la fibra de 10 nm hay un empaquetamiento del ADN de 5 a
7 veces. Este sin embargo, aún 1.000 veces menor que se produce en el cromosoma
metafásico
La Fibra de 30mn. Un Solenoide Formado por Nucleosomas.
Los
estudios de fibras de cromatina y de cromosomas con el microscopio electrónico
revelaron la presencia de una fibra ‘’Gruesa’’ diámetro varia entre 20 mn y 30
mn. Esta fibra corresponde probablemente ala estructura de una cromatina
inactiva se observa que la fibra de 30 mn esta formada por nucleosomas muy
compactos. Se origina por el plegamiento de la cadena de nucleosomas en una
estructura helicoida que tiene 6 nucleosomas por Vuelta.
Los Cromosomas Bacterianos No Forman Complejos con
Histonas.
Los
procationtes no poseen historia y los grupos fosfato libras en el ADN no
parecen ser neutralizados por proteínas básicas si no por otras sustancias,
probablemente poilaminas. Por otro lado, lados los organismo superiores
historias y en el curso de la evolución conservaron una gran medida su
secuencia de aminoácidos, lo que sugiere que estos tienen un papel esencial.
Estructura de la Cromatina
Los
estudios bioquímicos demostraron que los genes activos son mas accesibles a la
digestión de lo culceasa que las regiones del genomio inactivas. Esto puede
realizarse dirigiendo cucleos asilados con la encima ADNasa.En el critirocilo
los genes de globina y la región que los rodea son diez veces más accesibles
ala digestión que la masa de cromatina transcripcionalmente inactiva. El tejido
como el oviducho, en los que los genes de globina son inactivos, los genes no
son digeridos en forma preferencial. La cromatina de los genes activos se halla
menos condensados que la de la cromatinan inactiva que se encuentra quizás en la
forma de una fibra de 3mn. Otros componentes importantes de la cromatina son
algunas proteínas con histonicas abundantes denominadas HMG. Las HMG se
encuentran en cantidad elevada en la cromatina y una décima parte en todos los
nucleoso más de la célula está asociada como HMO. La mayor parte del ADN está
cubierta por nucleosomas los que no tienen especificidad de la secuencia de
Adn, Pero que contiene también signos de puntuaciones producidas por proteínas
que controlan los genes, firmemente unidas en sí.

FIGURA 5- IMAGEN 3D ESTRUCTURA DEL NÚCLEO INTERFÁSICO (MEMBRANAS).
LOS CROMOSOMAS
Durante
la división celular la cromatina es concentrada en los cromosomas. Después se
descubrimiento en 1876, los cromosomas
han sido objeto de estudio con el microscopio óptico y en 1910, Ya era evidente
que estaban relacionado con los fenómenos genéticos. Por esta razón el estudio
de los cromosomas es de mayor importancia. Analizamos aquí la morfología y la
composición de los cromosomas durante la mitosis.
CLASIFICACIÓN DE LOS CROMOSOMAS.
La
forma del cromosoma está determinada por
la posición del centrómero. Los cromosomas se pueden clasificar en cuatro
grupos de acuerdo con su forma, la que a su vez depende de la posición del centrómero (sitio donde se fijan la fibras del huso durante
la mitosis) que divide al cromosoma en sus dos brazos. El cromosoma telocéntrico presenta
el centrómero en uno de los extremos; el acrocentrico tiene un
brazo muy corto, el submetacéntrico tiene brazos de distinto largo,
y en el metacéntrico los brazos son iguales.

Durante el anafase los
cromosomas se doblan en la región del centrómero, de tal
manera que la metacéntrico aparece en forma de
"v", mientras que los acrocéntricos mantienen su
forma cilíndricas.

La citogenetistas crearon una compleja nomenclatura para describir los
componentes de los cromosomas, que seguramente el estudiante encontrara
confusa. Sin embargo, es importante recordar porque se refiere no solo alas características
morfológicas, sino también a propiedades específicas de los cromosomas.
Cromatina. En la metafase cada
cromosoma está formado por dos componentes simétricos, las cromatidas cada una
de las cuales contiene una sola molécula de ADN. Las cromatinas solo están unidas
entre sí a nivel del centometro y se separa al comienzo de la anafase, cuando
las cromatidas hermanas emigran a polos opuestos, por lo tanto, en la metafase
los cromosomas tiene dos cromatinas y en la anafase solo una.
Cromonema. Durante la profase (y a veces en la
interface) el material cromosómico se hace visible en forma de filameros finos,
llamados cromonemas, que presentan cromatidas en estados tempranos de condensación
por lo tanto las cromatida y cromonemas son dos hombres para una misma estructura.
Centrómero o cinetocoro. Es la región del
cromosoma donde convergen las fribias del huso mitofico se encuentra en una
parte más delgada del cromosoma la constricción primaria. Ambos lados del centrómero
hay secuencias de ADN muy repetitivo y
se tiñen más fuertemente con colorantes básicos (heterocromatina).
Los centrómeros contienen secuencias específicas de ADN y las que han sido
clonadas.
Algunas anormalidades cromosómicas causadas por ejemplo por los rayos X el
cromosoma puede perder el cinetocoro o presentar dos cinetocoro. Ambas aberraciones
son inestables
Telómero. Están denominación se refiere a los extremos
de los cromosomas. Los sitologos han reconocido por largo tiempo que los telómeros
tienen propiedades especiales. Si se fragmentan los cromosomas por rayos X los extremos
libres se tornan ¨pegajosos¨ y se fusionan con otros fragmentos y en cambio no
lo hacen con el telómero.
Satélite. Otro elemento morfológico de algunos
cromosomas es el satélite, que se presenta como un cuerpo estérico separado del
resto por una constricción secundaria, son característicos de cada cromosoma en
particular.
Constricción secundaria: son constantes en su posición
y tamaño y resultan útiles para identificar un cromosoma particular. Se distinguen
de las constricciones porque no hay una desviación angular de los segmentos cromosómicos.
Los organizadores nucleolares son ciertas constricciones secundarias en
las que los genes codifican a los ARN ribosómicos ARN.


Cada cromatina tiene una sola molécula de ADN.
Cada cromatida tiene una sola molécula lineal de ADN con sus proteínas asociadas.
Este concepto esencial para la concreción del comportamiento cromosómico,
fue denominado como teoría uninémica.
El ADN centromerico tiene secuencias reservadas.
Los cromosomas se alojan en sitios prefijados en el núcleo interfásico.
Es fácil comprender que los ciclos de condensación y descondensación que
ocurren durante la mitosis se facilitan si cada cromosomas ocupa un sitio
definido dentro del núcleo interfasico en células vegetales hay observaciones
que sugieren que en la interface los cromosomas tiene el mismo orden espacial
que tenían en la telofase eso se observó en la cebolla donde los cromosomas
tienen zonas heterocromaticas en los contromeros y telomeros. Es posible que
los cromosomas se dispongan en un orden fijo como lo sugiere el clásico trabajo
de Costelio.
HETEROCROMATINA
La heterocromatina corresponde
a zonas del cromosoma que permanecen condensadas durante la interface.
En 1928, Heitz la definió como
heterocromatina a las regiones del cromosoma que permanecen condensadas durante
la interface y proface temprana, formando los cromocentros o nucléolos falsos.
Las porciones restantes de los cromosomas, que permanecen en un estado no
condensado, fueron denominadas eucromatina. Heitz siguió a las células durante
el ciclo celular y encontró segmentos de cromosoma que no se descondesaban.
Los segmentos heterocromáticos tienden a localizarse en la
región pericentromérica de la mayoría de las plantas y animales, en los
telómeros, o en la vecindad de los organizadores nucleolares. En otros casos
todo el cromosoma se vuelve heterocromático.
Las zonas heterocromaticas
pueden visualizarse en los cromosomas condensados, porque se tiñen con mayor
fuerza o más débilmente que las regiones eucromáticas. Esto es lo que se
denomina heteropicnosis positiva o negativa de los cromosomas. Se piensa que en
la heterocromatina el ADN permanece fuertemente plegado en forma de fibras de
20-30nm, las cuales representan probablemente una configuración inactiva de la
cromatina.

FIGURA 9- FOTO DEL MICROSCOPIO HETEROCROMATINA
La heterocromatina puede ser
facultativa o constitutiva.
Heterocromatina constitutiva.-
Aparece condensada en todos los tipos celulares.
Heterocromatina facultativa.-
Solo se condensa en ciertos tipos celulares o en momentos especiales del
desarrollo. A menudo uno de los cromosomas se vuelve total o parcialmente
heterocromático.
La heterocromatina
constitutiva es la más común.
Este tipo de heterocromatina
contiene secuencias de ADN repetitivas que se denomina ADN satélite y puede
tener un rol estructural en los cromosomas.
La heterocromatina evidencia
varias propiedades que le son comunes, una de ellas es la replicación tardía.
La heterocromatina es generalmente inactiva desde el punto
de vista genético.
En general se acepta que la
cromatina condensada es inactiva en la síntesis de ARN.
Los cromosomas mitóticos
condensados no sintetizan ARN.
Hay evidencias que surgieren
que los segmentos heterocromáticos no se expresan genéticamente

FIGURA 10. Gato con coloración de manchas negras y
amarillas por lo cual se les denomina gatos tortuga. Son generalmente hembras.
Mary Lyon propuso que la pigmentación de manchas era producida por un gen
contenido en el cromosoma x. Este se vuelve heterocromático e inactivo en ciertas
células de la piel y no en otras.
La inactivación es al azar únicamente en células destinadas a dar origen al cuerpo. En cambio en células embrionarias que darán lugar a la placenta en los mamíferos siempre se inactiva el cromosoma x paterno.
Las hembras de otros mamíferos
y las mujeres no constituyen la excepción, su cuerpo representa un mosaico de
células, la mitad de las cueles tiene uno u otro cromosoma x en forma activa.
Estudios genéticos demostraron
que a los genes se les puede también inactivar si se les trasloca en regiones
heterocromáticas.
Es posible que el mecanismo de
inactivación de los genes en la cromatina condensada, sea más frecuente. En los
organismos multicelulares hay gran número de tejidos en los que ciertos genes
están activos, mientras que en otros están inactivos.
Parecería que el ADN ha
cambiado en los cromosomas x inactivos. Esto se deduce del hallazgo de que el
ADN, tomado de cromosomas x inactivos, no transforma las células de mamífero
que tienen una deficiencia en una enzima cuya actividad se detecta fácilmente y
que es codificada por el cromosoma x. En cambio el ADN extraído del cromosoma x
activo transforma las mismas células. El cambio más probable que afectaría al
propio ADN es la metilación.

FIGURA 11- EJEMPLO DE HETEROCROMATINA FACULTATIVA ES EL
MACHO PIANOCOCCUS CITRÍ, UN INSECTO EN EL CUAL TODO EL CONJUNTO DE CROMOSOMAS
PATERNOS ES HETEROCROMÁTICO. LOS GENES DE ESTOS CROMOSOMA SE HAN INACTIVO.
La
heterocromatina constitutiva tiene secuencias repetidas de ADN
La heterocromatina
constitutiva aparece en regiones del genomio que contienen cortas secuencias
repetitivas de ADN, denominadas ADN satélite.
El más conocido es el ADN satélite de ratón, una secuencia de 240 pares de bases que se repiten cerca de 1,000 000 de veces en el genomio de ratón, constituyendo el 10 % del ADN total de dicho
animal. E
Los
cromosomas y la heterocromatina
El estudio de los cromosomas es de la mayor importancia en
biología, ya que permite observar en
forma directa el comportamiento de las moléculas de ADN y de los genes.
Hay 4 tipos de cromosomas:
- · Teocéntricos
- · Acrocentricos
- Submetracentricos
- · metacéntricos
Cada cromosoma tiene dos
cromatidas, que están unidas a nivel del centrómero. Cada cromatida tiene una
sola molécula lineal de ADN. Se cree que los cromosomas tienen un armazón
central de proteínas no histonicas.
Las zonas heterocromicas
permanecen condensadas durante la interface y se tiñen en forma más intensa con
los colores básicos.
Se reconocen las heterocromatinas
faconstitutivas y facultativas
Las facontutivas pueden presentarse en la región Centroamérica y se relacionan con las secuencias repetitivas de ADN.
Las facultativas solo se condensan en ciertos
tipos celulares o en algunos estadios del desarrollo.
La iniciativa de los genes en la cromatina condensada es un
mecanismo que permite su regulación durante
la diferenciación.
martes, 28 de febrero de 2017
INTRODUCCIÓN A EL NÚCLEO INTERFÁSICO
Hola, bienvenidos.
El presente trabajo tiene como
finalidad dar a conocer; sobre el núcleo interfásico y partes que lo componen.
El núcleo interfásico:
Es la Parte elemental y el más voluminoso de la
célula eucariótica y se puede considerar como el “centro de control”, ya que en
él se localiza la mayoría del ADN celular que, como ya sabemos, determina todas
las funciones celulares.
Su constitución varía a lo largo de la vida celular,
distinguiéndose dos etapas; la interfase, durante la cual la
célula crece y se puede observar la forma y estructura del núcleo y
la división, durante la cual la membrana que rodea al núcleo desaparece,
con lo que los componentes del núcleo se mezclan con el citoplasma
El núcleo se pueden distinguir 2 estados morfológicos diferentes: el
núcleo interfásico y el núcleo en división.

FIGURA 1- FOTOMICROGRAFÍA DEL
NÚCLEO INTERFÁSICO ESTRUCTURA DE NÚCLEO INTERFÁSICO
LA
CROMATINA
La célula
del organismo eucarionte tiene un contenido de ADN constante que es característico
de cada especia y más grande que en las bacterias. Cuando el ADN se halla
empaquetado en un cromosoma metafísico debe plegarse entre 5000 y 10,000 veces.
En la cromatina el dan forman un complejo con las proteínas básicas (histonas).
Las historias son proteínas pequeñas con alto contenido en aminoácidos, arginina
y física. Hay cinco clases de Historia las H2A, H2B, H1 Y H4 se encuentran en
la proporción de dos moléculas por 200 pares de bases de ADN.

FIGURA 2- REPRESENTACIÓN
EN 3D DE HISTORIAS (HISTONE) EN LA CÉLULA Y EN EL ADN
La
cromatina está formada por unidades repetitivas, que en el microscopio
electrónico se presentan como esferas de 10 nm. En los extendidos aparecen a
modo de cuencas de collar. Cada nucleosoma contiene un octámero de historias
constituido por cada una de las cuatro historias. Alrededor del octámero hay
dos vueltas de ADN de 140 gb. La historia H se adhiere como puente de ADN que
une los nucleosomas y tiene 60 gb. Las cadenas de nucleosomas se pueden plegar
para formar la fibra ‘’gruesa’’ de 30 nm en el cual hay 6 nucleosomas por
vuelta en una hélice. Un empaquetamiento de ADN en el nucleosoma es de 5 a 7
veces, en la fibra gruesa de 40 veces y en el cromosoma metafísico de 5,0000 a
10,000.
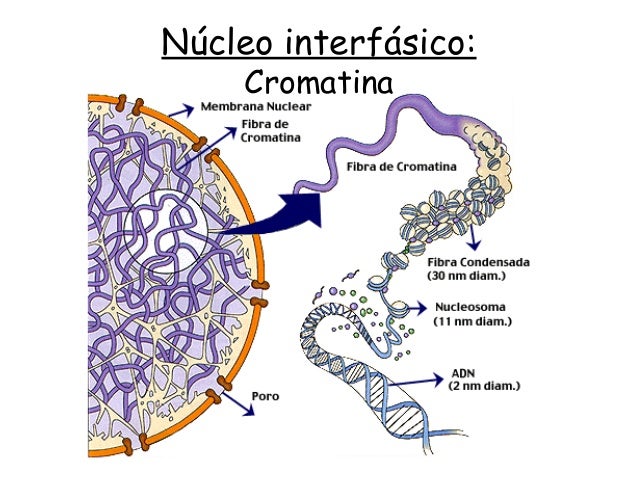
FIGURA 3- ESTRUCTURA DEL NÚCLEO INTERFÁSICO (CROMATINA).
LOS CROMOSOMAS
Durante
la división celular la cromatina es concentrada en los cromosomas. Después se descubrió en 1876, los cromosomas han sido objeto de
estudio con el microscopio óptico y en 1910, Ya era evidente que estaban
relacionado con los fenómenos genéticos. Por esta razón el estudio de los cromosomas
es de mayor importancia.
Los cromosomas se pueden clasificar en cuatro grupos de acuerdo con su forma, la que a su vez depende de la posición del centrómero (sitio donde se fijan la fibras del huso durante la mitosis) que divide al cromosoma en sus dos brazos. El cromosoma telocéntrico presenta el centrómero en uno de los extremos; el scrocentrimetro tiene un brazo muy corto, el submetacéntrico los brazos son iguales.

FIGURA 4- CLASIFICACIÓN DE LOS CROMOSOMAS.
Durante la anafase los cromosomas se doblan en la región del centrómero, de tal manera que la metacéntrico aparece en forma de "v", mientras que los acrocéntricos mantienen su forma cilíndricas.

FIGURA 4- REPRESENTACIÓN EN 3D DEL
CROMOSOMA Y EL ADN
Suscribirse a:
Comentarios (Atom)